
I didn’t expect to end up here.
When I began my career as an evolutionary biologist, I imagined fieldwork, maybe some molecular bench work, and a quiet academic life. I didn’t anticipate talking to geneticists, breeders, and data scientists about why their most frustrating obstacles aren’t about yield or resistance, but hidden deep inside the structure of plant genomes.

But here I am—because there’s something we’ve missed.
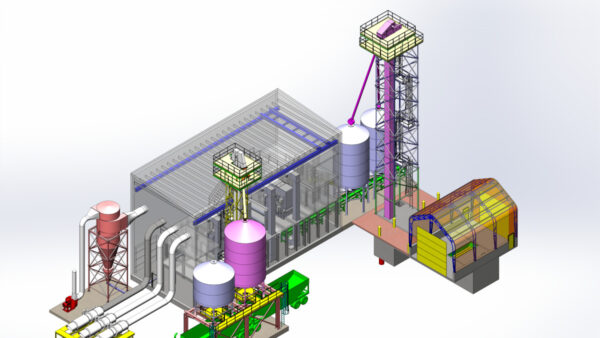
The real game-changer of the genomic era hasn’t been single nucleotide polymorphisms (SNPs). Sure, SNPs gave us fine-scale resolution and neat genome-wide association study (GWAS) signals. But the real beasts —the hidden forces steering everything from trait inheritance to hybrid sterility — are structural variants.
I’m talking about deletions, duplications, inversions and translocations.
These are not minor anomalies. They’re massive. They affect far more of the genome than SNPs ever will. And they’re not just common — they’re consequential.
Sunflowers, as it turns out, are the perfect model for this structural complexity, even though they’re terrible in many other ways. They’re large, hard to transform, and slow to work with. But their genome? It’s like a structural variation theme park.
When we sequenced 10 cultivated sunflower genomes, we found ~12 million SNPs. Not bad. But alongside those were ~40,000 copy number variants, 10,000 translocations, and several hundred inversions. This isn’t just messy. It’s foundationally different from what we expected.
And it matters.
When breeders try to bring in traits from wild relatives—say, resistance to broomrape or Sclerotinia — they often run into a wall. That wall isn’t the trait itself. It’s the structural rearrangements. These changes suppress recombination and make it nearly impossible to cleanly integrate new alleles without dragging along a mess of unwanted genetic baggage. Worse, some rearrangements trigger hybrid sterility outright.
Take Helianthus debilis, a close sunflower cousin often tapped for introgression. It differs from cultivated sunflower by eight large translocations. Try crossing the two, and you’ll find yourself fighting suppressed recombination and plummeting seed viability.
It raises a big evolutionary question: If structural variants reduce fitness, how do they become established? We used to think this was due to chance (i.e., genetic drift). Now, we suspect something else— female meiotic drive, in which selfish genetic elements compete for inclusion in the egg, with the structural variants along for the ride. Either way, they’re not going anywhere.
Among all SVs, inversions are the most insidious — and the most fascinating. Because they don’t always harm fertility, they tend to go undetected. Yet they quietly suppress recombination across vast genomic regions, effectively locking together adaptive alleles into what we call supergenes.
It sounds elegant—nature bundling useful traits into single, inheritable blocks. But those bundles often come with a catch. One inversion might boost seed size, sure — but it may also reduce oil content or delay flowering. That’s the double-edged sword breeders live with.
This isn’t just a sunflower story. Similar patterns are now emerging in maize, barley and more. Structural variation is turning out to be a near-universal language for local adaptation in plants.
So, what’s the breeding takeaway?
Yes, you can harness these supergenes for stress tolerance and climate adaptation. But you have to understand the trade-offs.
So where does this leave us?
Structural variation isn’t a nuisance to be managed — it’s the architecture of evolution. These rearrangements define how traits move (or don’t move) through breeding populations. They create opportunity and constraint, simultaneously.
We can’t ignore them anymore.
If we want to future-proof our crops for climate, disease, and demand, we need to build tools and strategies that respect the structural architecture of the genome. That means sequencing more pangenomes, mapping SVs across populations, and using gene editing to decouple trait trade-offs, and designing breeding programs that recognize when the roadblock isn’t the gene — it’s the context it lives in.
The genomic revolution gave us sharper questions. The answers will decide the future of breeding — and whether we’re ready to work with nature’s blueprints, instead of against them.